

Home
BELEMNITEN DER OBERKREIDE
von Carsten Rohde
Die Familie Belemnitellidae PAVLOW, 1914
|
Teil 3:
Artbegriff und Artenbestimmung |
Teil 3: Artbegriff und Artenbestimmung
Jede Artbestimmung ist zunächst eine Frage des subjektiven optischen Eindrucks. Sie ist nie wahr, sondern stets nur wahrscheinlich. Die verbale Beschreibung eines Rostrums ist durch die Wahl und die individuelle Interpretation der verwendeten, das Fossil beschreibenden Adjektive, wie klein/groß, kegel-/pfahl-/pflockförmig/zylindrisch oder etwa schlank/gedrungen, nicht widerspruchsfrei. Die Übergänge sind vielseitig und die Schattierungen zahlreich. So bezeichnet Jurij Alexander Jeletzky (1951, S.78f) die Rostrumform sowie die Ausprägung oder die Stärke der Rostrumskulptur der Belemnitellen und Belemnellen als zur Bestimmung vollkommen unzuverlässig. Deshalb ist bei den merkmalsarmen Resten der Belemnitelliden eine Hinwendung zur Inanspruchnahme der "inneren" Merkmale (Jugendrostren, Alveolentiefe, Apikallinie) sowie biometrischer Verhältniszahlen der gemessenen Merkmalswerte zu erkennen, die mit statistisch empirischen Methoden gewonnen, untersucht und, wie in nachstehender Tabelle 1 von Walter Kegel Christensen (2000, S. 60) dargestellt werden. Mittelwerte der gewählten Eigenschaften von betrachteten Populationen bleiben jedoch abstrakt. Nur das einzelne Individuum, das vermessene Rostrum ist real.
Tabelle 1 + Abbildung 1
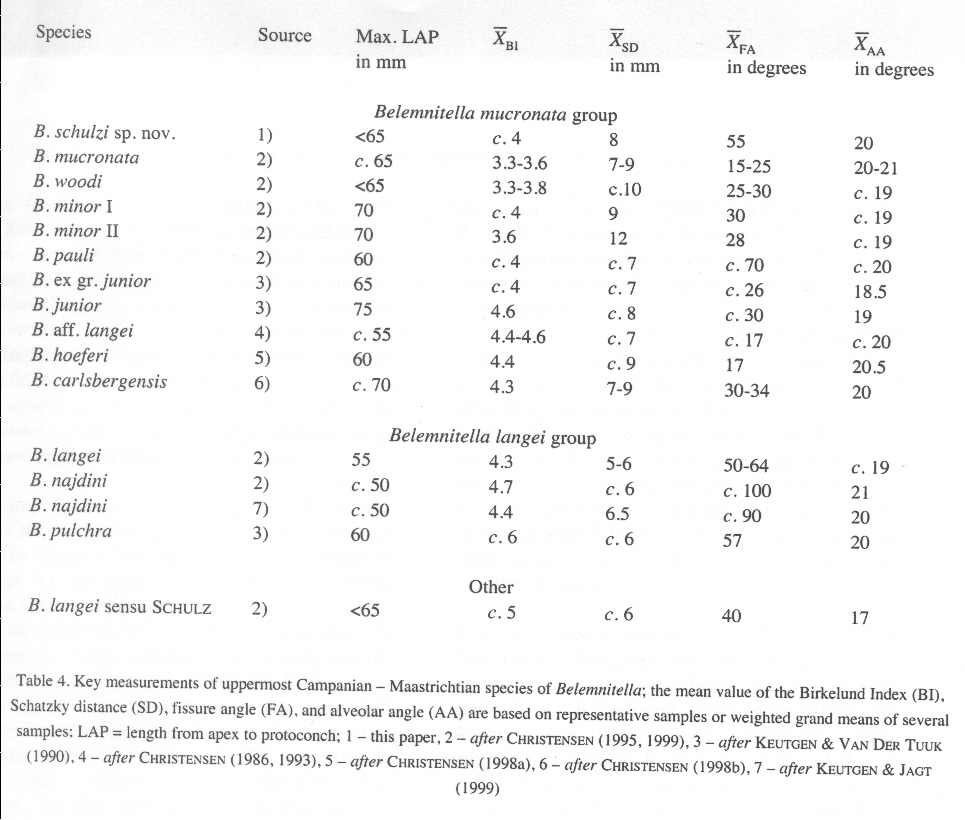
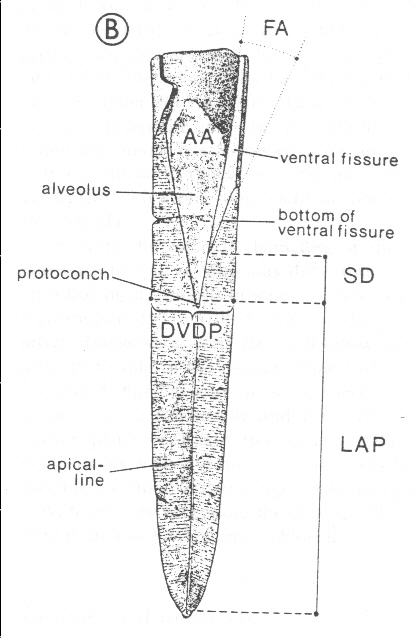 (LAP = Länge vom Apex bis zum Protoconch; BI = Birkelund-Index = LAP/Rostrumdicke in der Schlitzebene in Höhe des Protoconchs DVDP; SD = Schatzky-Wert = Entfernung vom Protoconch bis zur Einmündung des Ventralschlitzes in die Alveole; AA = Alveolarwinkel)
(LAP = Länge vom Apex bis zum Protoconch; BI = Birkelund-Index = LAP/Rostrumdicke in der Schlitzebene in Höhe des Protoconchs DVDP; SD = Schatzky-Wert = Entfernung vom Protoconch bis zur Einmündung des Ventralschlitzes in die Alveole; AA = Alveolarwinkel)
Die Wahl von Merkmalen führt immer zu typologischen Arten, sogenannten Morphospezies, die, im Gegensatz zu biologischen Arten, jederzeit aufgelöst und neu gegliedert werden können. Der paläontologische Artbegriff ist praktisch deshalb gleich dem morphologischen Artbegriff, da an Fossilien artkennzeichnende, biologische Merkmale zumeist nicht erhalten sind und die im Zeitquerschnitt geltende reproduktive Isolation nicht nachgewiesen werden kann (- wie kreuzt man zwei Rostren?). Ergebnis dieses Dilemmas: Jedwede vom jeweiligen Forscher als signifikant eingeschätzte Formabweichung erhält den Status eines artbestimmenden Merkmals; und im Zeitlängsschnitt werden Populationen aufeinander folgender geologischer Schichten solange als einer Art zugehörig betrachtet (einer sog. Chronospezies), wie ihre Merkmale nicht über einen statistisch als relevant betrachteten Bereich der empirisch erhobenen Merkmalswerte hinausreichen. Doch Veränderungen der Umwelt in der Zeit können das sichtbare Erscheinungsbild (den Phänotyp) folgender Generationen von Populationen beliebig stark wandeln, ohne zwangsläufig zur Umwandlung einer Art in eine andere neue Art (ein Wandel des Genotyps) zu führen, sondern lediglich zum Wechsel in den Merkmalsunterschieden zeitlich gestaffelter Populationen ein und derselben Spezies. Werden diese, durch die phänotypische Plastizität hervorgerufenen "Evolutions"stufen einer lückenlosen, sich nicht aufspaltenden Formenreihe als verschiedene Arten bezeichnet, so ist zunächst festzustellen, dass hierfür keine natürlichen Grenzen existieren. Diese Grenzen werden erst durch subjektive Gliederung der Formenreihe mittels empirischer Statistik geschaffen, sind damit im Sinne des biologischen Artkonzeptes willkürlich und definieren allenfalls mögliche, häufig auch sinnvolle Abgrenzungen auf dem Niveau von Unterarten.
Als eine gelungene zeitliche Aufspaltung einer phylogenetischen Entwicklungsreihe wird die Belemnitengattung Gonioteuthis BAYLE 1878 betrachtet. Ausgehend von Gonioteuthis praewestfalica ERNST & SCHULZ 1974 im späten Mittel-Coniac, über Gonioteuthis westfalica SCHLÜTER 1874 erfolgt eine lückenlose Entwicklungsreihe zu Gt. westfalicagranulata STOLLEY 1897 , Gt. granulata BLAINVILLE 1827, Gt. granulataquadrata STOLLEY 1897, Gt. quadrata quadrata BLAINVILLE 1827 bis zu Gt. quadrata gracilis STOLLEY 1892 im Oberen Unter-Campan. Die Abgrenzung der Gonioteuthisarten erfolgt anhand des Riedel-Quotienten, eines biometrischen Wertes, gebildet als Verhältnis von Rostrumlänge zu Alveolentiefe. Die Forschungen von Gundolf Ernst (1964, S. 169) zeigen Etappen der Stammesgeschichte der Gonioteuthiden mit allmählichen Übergängen und sich überschneidenden Abschnitten der Evolutionsprofile - Indizien für eine lückenlose, nicht verzweigte phylogenetische Entwicklung der Gattung Gonioteuthis. Mithin liegt der Verdacht nahe, dass es sich bei den Arten der Gattung Gonioteuthis um Chronospezies handelt. In Anwendung des biologischen Artbegriffs müssten daher alle Gonioteuthisarten als Unterarten eines polytypischen Stammes angesehen werden, der nach Blainville 1827 entweder mit Gonioteuthis granulata oder quadrata zu benennen wäre, zuzüglich eines differenzierenden Namens für die jeweilige Unterart.
Tabelle 2
Die Aufgabe empirischer Wissenschaft besteht darin, befriedigende Erklärungen zu finden, die nicht aus dem Augenblick heraus entstanden sind und überprüft werden können. Empirische Wissenschaft basiert dabei stets auf Stichproben mit endlicher Zahl von Individuen. Je größer die betrachtete Population, desto aussagefähiger die gewonnenen Ergebnisse. Stichproben mit nur wenigen Individuen können zu dubiosen Ergebnissen führen, zur Festlegung von Arten mit geringer Aussagefähigkeit und Akzeptanz.
Nach W. K. Christensen (2000,S.62) wurde die Belemnitellidenart Belemnitella posterior KONGIEL 1962 anhand von 7 nahezu vollständigen und 3 fragmentarischen Rostren im Übergang Obercampan/Untermaastricht begründet. Die mittleren statistischen Werte der von Christensen vermessenen Rostren liegen, wie nachstehende Tabelle zeigt, im selben Bereich wie die Werte der von Christensen neu etablierten Art Belemnitella schulzi CHRISTENSEN 2000 und sind der Art Belemnitella minor II CHRISTENSEN so ähnlich, dass er selbst "noted that it was uncertain if these were extreme variants of B. minor II or represented a separate species."
Da sich auch die stratigrafischen Reichweiten der vorgenannten Arten überlappen, erscheint eine Identität von posterior und schulzi nicht ausgeschlossen, wobei posterior der ältere Artname wäre. Und was hindert uns daran den Bogen größer zu schlagen und den Zweifeln Christensens folgend beide vorgenannten Arten als Varianten in Belemnitella minor II zu integrieren?
Doch auch bei hinreichend großer Zahl der untersuchten Rostren: Berechnungen anhand von Stichproben verschaffen keine Gewissheit, sondern nur Wahrscheinlichkeit. So sind stark vom Mittelwert abweichende Ergebnisse in der Paläontologie nicht zwingend Ausdruck von Pathologie und damit zu eliminieren; Abweichler können auch Ausdruck einer hohen Variabilität der Art sein, d.h. auf ein großes Potenzial an Variantenbildung hinweisen. Sie können aber auch schlicht eine neue Art angeben. Doch wo hört die Variabilität einer Art, ihre phänotypische Plastizität auf, wo wird die Normalität der einen Art zu der Normalität ihrer Schwesterart? Es gibt innerhalb der Belemnitengattungen durchaus Arten hoher stratigraphischer Reichweite und grosser Variabilität , wie Hibolites jaculoides SWINNERTON 1936 (Ober-Valangin Oberstes Barrême), neben solchen mit kurzer Lebensdauer und unveränderlichen Merkmalsausprägungen (Belemnellocamax boweri CRICK 1910, frühes Mittel-Cenoman).
Weitere Schwierigkeiten bei der Artabgrenzung ergeben sich aus den Problemen der Formähnlichkeit (Homöomorphie, so tauchen in der Entwicklungsgeschichte der Belemniten zu verschiedenen Zeiten kleine, keulenförmige Rostren auf, die kaum zu unterscheiden sind), der Heterogenität zwischen Individuen unterschiedlichen Alters, durch ausgeprägten Geschlechtsdimorphismus (von Schulz bei den Belemnellen angenommen, von Peter Doyle bei Youngibelus aus dem Toarc von Yorkshire beschrieben) und pathologische Veränderungen an Rostren gleicher Artzugehörigkeit. Zudem sind die Individuen einer Art in ihren Merkmalsausprägungen nie deckungsgleich; wie auch Populationen gleicher Art an verschiedenen Standorten, hervorgerufen durch die abweichenden Milieubedingungen, möglicherweise erhebliche Merkmalsunterschiede aufweisen werden.
Bei der Bestimmung von Arten der Belemnitenfamilie Belemnitellidae werden verschiedene biometrische Verhältniszahlen (Riedel-Quotient, Birkelund-Index) oder gar von komplexen Merkmalskombinationen (bei der Abgrenzung der Belemnellen) gebildet. Derartige Verhältniszahlen sind aber nur dann geeignet Arten zu differenzieren, wenn ihre Schwankungen nur Ausdruck der Normabweichung sind und nicht durch Messfehler oder unterschiedliches Wachstumsverhalten im Alterungsprozess beeinflusst wird.
Der gezielte Einsatz der Variationsstatistik und biometrischer Messmethoden bei der Bestimmung von Belemnitenrostren kann als Mittel dienen, eine Auflösung des Artbegriffs durch unterschiedliche Interpretation beschreibender, qualitativer Merkmale zu verhindern. Sie kann aber auch das Gegenteil bewirken, da sie nicht wie die altmodische subjektive Deutung von der Gesamtheit der beobachteten Merkmale ausgeht, sondern nur von einigen biometrisch erfassbaren. Auch sollte die Variabilität der Arten nicht zu gering geachtet werden, wie viele Beispiele aus der Neontologie zeigen.
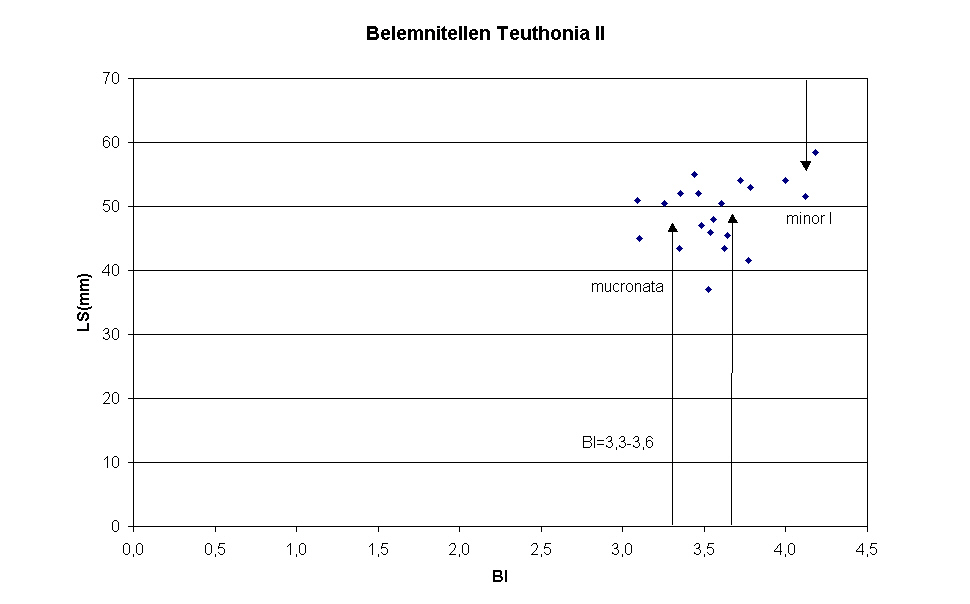
Im Jahre 2005 wurden in der Grube Teutonia Nord der Teutonia AG in Misburg (bei Hannover), die stratigraphisch von der pilula/senonensis-Zone des Unter-Campan bis zur minor/polyplocum-Zone des Unteren Ober-Campan reicht, 20 Belemnitenrostren nicht horizontiert gesammelt. Nach Reinigung der Rostren mit Hilfe des Tensids Rewoquat®, wurden anhand der Alveole geprüft, ob die Rostren zu der Gattungen Gonioteuthis BAYLE 1879 (kurze Pseudoalveole mit wenige mm langem Alveolarschlitz und wulstigem Alveolarrand) oder Belemnitella dORBIGNY 1840 (ausgeprägte Alveole und langer Alveolarschlitz) gehören. Alle Rostren konnten der Gattung Belemnitella zugeordnet werden. Zur Bestimmung der Lage des Protoconch wurde die Alveole, soweit möglich, aufgebohrt; bei den meisten Rostren war indes das eingeschlossene Sediment zu fest, um es zu entfernen. Diese Exemplare wurden in Höhe der vermuteten Lage der Embryonalkammer quergeteilt. Aufgrund der nunmehr geringeren Alveolentiefe konnte die Position des Protoconchs durch Entfernen des geringen verbliebenen Sediments leicht bestimmt werden. Lag der Bruch Apex-seitig unterhalb der Embryonalkammer, wurde das alveolenseitige Rostrumfragment in der Alveolarschlitzebene gespalten. Die Werte für die Rostrumlänge bis zu Protoconch LAP sowie der Birkelund-Index BI wurden in Diagramm 1 abgetragen. Das Schaubild zeigt, dass der Birkelund-Index der meisten Rostren innerhalb des von Christensen in Tabelle 1 angegebenen Bereichs von 3,3 bis 3,6 liegt, der Belemnitella mucronata LINK 1807 repräsentiert. Die BI- Werte der Rostren 1, 2 und 12 entsprechen dem Mittelwert des BI-Index für Belemnitella minor JELETZKY 1951 in der von Christensen eingeführten Variante minor I.
|
Analyse III:
Belemnitella Arten aus der Grundgesamtheit von 20 nicht Horizont orientiert gesammelten Rostren in Teutonia Nord/Misburg
|
|||
|
Rostrum
|
LS(LAP)
|
DP
|
BI
|
| 1 | 54 | 13,5 | 4,0 |
| 2 | 58,5 | 14 | 4,2 |
| 3 | 53 | 14 | 3,8 |
| 4 | 46 | 13 | 3,5 |
| 5 | 45 | 14,5 | 3,1 |
| 6 | 51 | 16,5 | 3,1 |
| 7 | 45,5 | 12,5 | 3,6 |
| 8 | 41,5 | 11 | 3,8 |
| 9 | 43,5 | 12 | 3,6 |
| 10 | 47 | 13,5 | 3,5 |
| 11 | 52 | 15 | 3,5 |
| 12 | 51,5 | 12,5 | 4,1 |
| 13 | 54 | 14,5 | 3,7 |
| 14 | 50,5 | 14 | 3,6 |
| 15 | 48 | 13,5 | 3,6 |
| 16 | 50,5 | 15,5 | 3,3 |
| 17 | 55 | 16 | 3,4 |
| 18 | 52 | 15,5 | 3,4 |
| 19 | 37 | 10,5 | 3,5 |
| 20 | 43,5 | 13 | 3,3 |
| 21 | 51 | 15,5 | 3,3 |
| 22 | 53 | 14 | 3,8 |
|
STABW
|
0,3
|
||
|
Mittelwert
|
3,6
|
||
Literaturhinweise:
Christensen, W.K. (1997): The Late Cretaceous belemnite family Belemnitellidae: Taxonomy and evolutionary history. Bulletin of the Geol. Soc. Denmark, Vol. 44, S. 59-88, Kopenhagen.
Christensen, W.K. (2000): Belemnitella schulzi sp.nov. from the uppermost Campanian and lowest Maastrichtian chalks of northwest Germany and Denmark. Acta Geologica Polonica, Vol. 50, S. 55 66, Warschau.
Ernst, G. (1964): Ontogenie, Phylogenie und Stratigraphie der Belemnitengattung Gonioteuthis BAYLE aus dem nordwestdeutschen Santon/Campan. Fortschr. Geol. Rheinld. u. Westf. 7, Krefeld.
Jeletzky, J.A. (1951): Die Stratigraphie und Belemnitenfauna des Obercampan und Maastricht Westfalens, Nordwestdeutschlands und Dänemarks sowie einige allgemeine Gliederungs-Probleme der jüngeren borealen Oberkreide Eurasiens. Beih.Geol.Jb. 1, Hannover.
Mayr. E. (1975): Grundlagen der zoologischen Systematik, Hamburg/Berlin.
Schulz, M.-G. (1979). Morphometrisch-variationsstatistische Untersuchungen zur Phylogenie der Belemnitengattung Belemnella im Untermaastricht NW-Europas, Geol.Jb. A47, Hannover.
|
Teil 3:
Artbegriff und Artenbestimmung |