

Home
"Die Ostsee" - Vergangenheit, Gegenwart und Zukunft
von Klaus Vöge
 Die Ostsee ist eines der bevorzugten Urlaubsgebiete der Deutschen. Doch abseits der idyllischen Sandstrände und Steilküsten ist die Ostsee ein relativ junges Binnenmeer, das mit erheblichen Umweltproblemen zu kämpfen hat. Mit eben 415.000 km² und einem Wasservolumen von 21.600 km³ zählt die Ostsee zu den kleinen Meeren unseres Planeten. Da durch die schmale Verbindung zur Nordsee das Wasser nur langsam ablaufen kann, ist der Pegelstand im Nordosten 35 cm höher als in der westlichen Ostsee. Durch die Flusswasserzuläufe im östlichen Bereich, kommt es von Westen nach Osten zur Aussüßung. Im Gebiet Dänemarks liegt der Salzgehalt zwischen 2 bis 3%, während er zum östlichen Teil kaum noch spürbar ist.
Die Ostsee ist eines der bevorzugten Urlaubsgebiete der Deutschen. Doch abseits der idyllischen Sandstrände und Steilküsten ist die Ostsee ein relativ junges Binnenmeer, das mit erheblichen Umweltproblemen zu kämpfen hat. Mit eben 415.000 km² und einem Wasservolumen von 21.600 km³ zählt die Ostsee zu den kleinen Meeren unseres Planeten. Da durch die schmale Verbindung zur Nordsee das Wasser nur langsam ablaufen kann, ist der Pegelstand im Nordosten 35 cm höher als in der westlichen Ostsee. Durch die Flusswasserzuläufe im östlichen Bereich, kommt es von Westen nach Osten zur Aussüßung. Im Gebiet Dänemarks liegt der Salzgehalt zwischen 2 bis 3%, während er zum östlichen Teil kaum noch spürbar ist.
 Vergangenheit: Während die Nordsee auf eine Geschichte von 180 Millionen Jahren zurückblicken kann, liegt der Ursprung der Ostsee im Dunkeln bzw. unter Eis vergraben. Wie die Elster- und Saaleeiszeit an der Bildung der Ur-Ostsee beteiligt waren, ist nicht geklärt. Die Saalevereisung schuf jedoch vor ca. 100.000 Jahren die Barriere zwischen Nord- und Ostsee. Die damaligen Endmoränen der Geest von Schleswig-Holstein und Jütland sind von der folgenden Weichsel-Vereisung nicht erreicht worden und somit erhalten geblieben. Gesichert ist, dass bei der Formung der Ostsee-Senke die Ausschürfungen der Weichsel-Kaltzeit eine wesentliche Rolle gespielt haben. Die größte Ausdehnung der Weichseleiszeit war vor 45.000 Jahren und reichte bis an die Linie Flensburg – Rendsburg – Neumünster – Hamburg – Berlin. Auf dem skandinavischen Schild lag derzeit eine Eisdecke von 3.000 bis 5.000 m. Selbst in Schleswig-Holstein hatte das Eis immer noch eine Stärke von 200 bis 400 m. Dieses bedeutete, dass auf die auf dem Magma schwimmende Erdkruste ein enormer Druck ausgeübt wurde. Aus der Erkenntnis, dass die Erdkruste nicht starr ist, kommt es bei lang anhaltender Belastung zum Einbiegen/Senken der Erdkruste. Bei Entlastung wölbt sich die Erdkruste aufgrund der Zähigkeit des Magmas etwas verzögert wieder auf, so dass es mit dem Abschmelzen des weichseleiszeitlichen Inlandeises zu einer Ausgleichsbewegung kam. Diese Hebung der Erdkruste ist bis zum heutigen Tag noch nicht abgeschlossen.
Vergangenheit: Während die Nordsee auf eine Geschichte von 180 Millionen Jahren zurückblicken kann, liegt der Ursprung der Ostsee im Dunkeln bzw. unter Eis vergraben. Wie die Elster- und Saaleeiszeit an der Bildung der Ur-Ostsee beteiligt waren, ist nicht geklärt. Die Saalevereisung schuf jedoch vor ca. 100.000 Jahren die Barriere zwischen Nord- und Ostsee. Die damaligen Endmoränen der Geest von Schleswig-Holstein und Jütland sind von der folgenden Weichsel-Vereisung nicht erreicht worden und somit erhalten geblieben. Gesichert ist, dass bei der Formung der Ostsee-Senke die Ausschürfungen der Weichsel-Kaltzeit eine wesentliche Rolle gespielt haben. Die größte Ausdehnung der Weichseleiszeit war vor 45.000 Jahren und reichte bis an die Linie Flensburg – Rendsburg – Neumünster – Hamburg – Berlin. Auf dem skandinavischen Schild lag derzeit eine Eisdecke von 3.000 bis 5.000 m. Selbst in Schleswig-Holstein hatte das Eis immer noch eine Stärke von 200 bis 400 m. Dieses bedeutete, dass auf die auf dem Magma schwimmende Erdkruste ein enormer Druck ausgeübt wurde. Aus der Erkenntnis, dass die Erdkruste nicht starr ist, kommt es bei lang anhaltender Belastung zum Einbiegen/Senken der Erdkruste. Bei Entlastung wölbt sich die Erdkruste aufgrund der Zähigkeit des Magmas etwas verzögert wieder auf, so dass es mit dem Abschmelzen des weichseleiszeitlichen Inlandeises zu einer Ausgleichsbewegung kam. Diese Hebung der Erdkruste ist bis zum heutigen Tag noch nicht abgeschlossen.
Der zweite wichtige Einflussfaktor bei der Entwicklung der Ostsee sind die klimaabhängigen Wasserstandsänderungen. Da zur damaligen Zeit große Wassermassen im Eis gebunden waren, lag der Wasserspiegel, zum Beispiel in der Nordsee, um etwa 80 bis 100 Meter unter dem heutigen Niveau. Das änderte sich erst, als es vor ca. 15.000 Jahren im Zuge der einsetzenden Warmphase zur Freisetzung großer Wassermassen kam. Das Zusammenspiel von Landhebung, weltweitem Meeresspiegelanstieg und unterschiedlichem Salzgehalt bestimmte entscheidend die spät- und nacheiszeitliche Geschichte der Ostsee. Im späten Pleistozän bildeten sich vor dem Eisrand in Südschweden etliche Eisstauseen, die sich in der jüngeren Tundrenzeit zum großen „Baltischen Eisstausee“ (einem Süßwassersee) zusammenschlossen. Der Stausee füllte sich weiter, bis er 25 bis 30m über dem Nordseewasserstand lag und mit einem katastrophen-artigen Ereignis überlief. Riesige Mengen von Süßwasser ergossen sich durch Mittelschweden direkt in den Skagerrak. Durch den entstandenen mittelschwedischen Verbindungskanal konnte nun auch Salzwasser in den Ostseebereich einfließen. So bildete sich das brackige „Yoldia-Meer“, das nach einer damals dort lebenden Brackwasser-Muschel benannt ist.
Dieser frühen „Ostsee“ war nur eine kurze Zeit vergönnt. Mit dem rasch abschmelzenden Eis stieg der weltweite Meeresspiegel, doch hob sich auch das vom Gewicht des Eises befreite Mittelschweden. Die Verbindung zum Skagerrak schloss sich weitgehend und es bildete sich der „Ancylus-See“ (ein Süßwassersee). Auch hier wurde der See nach der dort lebenden Flussnapfschnecke genannt. Mit dem weiter steigenden weltweiten Meeresspiegel war auch die zweite Süßwasserphase bald vorüber. Unaufhörlich drang Salzwasser durch den Kattegatt nach Süden vor. Es schufen sich schließlich durch den Öresund sowie den Großen und Kleinen Belt drei Verbindungswege zur Ostsee. Mit dem „Litorina-Meer“ wurde die wechselvolle Süß- und Salzwasser-Geschichte dieses Schelfmeeres vorerst beendet und es entstand die Brackwasser-dominierte Ostsee, wie wir sie heute kennen. In den letzten 2.000 Jahren, süßte die Ostsee wegen der ständigen Zufuhr von Flusswasser immer mehr aus. Zudem kam es aufgrund der Verengung der dänischen Pforten zu einem verringerten Salzwassereinstrom. Der ältere Abschnitt dieser Epoche wird nach der Brackwasserschnecke Limnea ovata als „Limnea-Meer“ bezeichnet; die letzten 500 Jahre werden nach der brackigen Sandklaffmuschel Mya arenaria als „Mya-Meer“ benannt.
(Bilder anklicken, um die Galerie zu öffnen.)



")

")

")
")
Gegenwart: Zu den Besonderheiten der Ostsee gehört, dass es nur über die Verbindung zur Nordsee einen eingeschränkten Wasseraustausch mit den salzhaltigen Weltmeeren gibt. Der Salzwassereinstrom aus dem Westen trifft auf gewaltige Süßwassermengen, die über große Flüsse wie Oder, Weichsel, Memel, Düna und Newa in die Ostsee gelangen. Dieses Gemisch aus Süß- und Salzwasser macht die Ostsee zu einem der größten Brackwassergebiete der Erde. Da die Salzkonzentration nicht einheitlich verteilt ist, sondern in der westlichen Ostsee deutlich höher ist als in den östlichen und nördlichen Regionen, steht das Ökosystem unter Dauerstress. Viele Organismen haben erhebliche Schwierigkeiten mit dem unterschiedlichen Salzgehalt. Die Ostsee ist eine geschundene Region, manche meinen sogar, sie sei auf dem Weg, ein totes Meer zu werden. Pflanzen und Tiere finden hier keine optimalen Lebensbedingungen vor, was nicht zuletzt am wechselhaften Nährstoffkreislauf liegt. Überdüngung des Meeres führt vor allem in den Sommermonaten zu einem vermehrten Algenwachstum. Wenn diese absterben, sinken sie auf den Meeresboden, wo sie von Bakterien zersetzt werden. Bei diesem Vorgang wird dem am Boden lagernden Salzwasser Sauerstoff entzogen. Zusätzlich setzen die Bakterien Schwefelwasserstoff frei, der für Fische giftig ist. Wenn sauerstoffhaltiges Nordseewasser für längere Zeit ausbleibt, stirbt der Meeresboden. Das muss allerdings nicht so bleiben. Diese außergewöhnliche Salzzufuhr geschah 2014.
 Das Leibniz Institut Warnemünde meldet: Im Dezember 2014 kam es in der Ostsee zu einem für ihr Ökosystem wichtigen, aber sehr seltenen Phänomen: Sauerstoffhaltiges Salzwasser strömte über mehrere Tage aus der Nordsee in die Ostsee ein. Nach Auswertung der gemessenen Werte steht fest, dass es sich um den größten Salzwasser-Einbruch der letzten 60 Jahre handelte. Innerhalb von drei Wochen gelangten 198 km³ sauerstoffhaltiges Salzwasser (Das entspricht etwa der Wassermenge im Bodensee) in die Ostsee. Das bedeutet ca. 4 Gigatonnen Salz. Vorausgegangen war eine Phase langanhaltender Ostwinde, die einen starken Ausstrom verursachten, sodass der Meeresspiegel deutlich fiel. Als dann der Wind nach West drehte, konnte das Nordseewasser einströmen. Da die Windsituation über drei Wochen hinweg unverändert blieb, konnten große Mengen an Salzwasser in die Ostsee gelangen. Etwas Besseres konnte der Ostsee zurzeit nicht passieren.
Das Leibniz Institut Warnemünde meldet: Im Dezember 2014 kam es in der Ostsee zu einem für ihr Ökosystem wichtigen, aber sehr seltenen Phänomen: Sauerstoffhaltiges Salzwasser strömte über mehrere Tage aus der Nordsee in die Ostsee ein. Nach Auswertung der gemessenen Werte steht fest, dass es sich um den größten Salzwasser-Einbruch der letzten 60 Jahre handelte. Innerhalb von drei Wochen gelangten 198 km³ sauerstoffhaltiges Salzwasser (Das entspricht etwa der Wassermenge im Bodensee) in die Ostsee. Das bedeutet ca. 4 Gigatonnen Salz. Vorausgegangen war eine Phase langanhaltender Ostwinde, die einen starken Ausstrom verursachten, sodass der Meeresspiegel deutlich fiel. Als dann der Wind nach West drehte, konnte das Nordseewasser einströmen. Da die Windsituation über drei Wochen hinweg unverändert blieb, konnten große Mengen an Salzwasser in die Ostsee gelangen. Etwas Besseres konnte der Ostsee zurzeit nicht passieren.
 Zukunft: Wie sieht die Zukunft der Ostsee aus? Die globale Klimaerwärmung wird auch hier nicht folgenlos bleiben; durch das Abschmelzen polarer Eiskappen steigt der Meeresspiegel. Dennoch sind entlang der Ostseeküsten weit weniger Flächen von künftigen Überschwemmungen bedroht als zum Beispiel rings um die Nordsee. Die Hügelketten der eiszeitlichen Moränen schützen fast überall das Hinterland. Nur in der Nähe der Flussmündungen von Oder, Weichsel und Memel sind größere Flächen gefährdet. In Schleswig-Holstein sind vor allem die nördliche Probstei, der Oldenburger Graben und Teile von Fehmarn bedroht. Auch wenn das beruhigend klingen mag, sollten wir uns als Verursacher des Klimawandels über die Gefahren für viele andere Küsten auf der Welt und die dort wohnenden Menschen im Klaren sein und entsprechend verantwortungsbewusst handeln. Nur dann können wir mit gutem Gewissen die Schönheit der Ostsee genießen.
Zukunft: Wie sieht die Zukunft der Ostsee aus? Die globale Klimaerwärmung wird auch hier nicht folgenlos bleiben; durch das Abschmelzen polarer Eiskappen steigt der Meeresspiegel. Dennoch sind entlang der Ostseeküsten weit weniger Flächen von künftigen Überschwemmungen bedroht als zum Beispiel rings um die Nordsee. Die Hügelketten der eiszeitlichen Moränen schützen fast überall das Hinterland. Nur in der Nähe der Flussmündungen von Oder, Weichsel und Memel sind größere Flächen gefährdet. In Schleswig-Holstein sind vor allem die nördliche Probstei, der Oldenburger Graben und Teile von Fehmarn bedroht. Auch wenn das beruhigend klingen mag, sollten wir uns als Verursacher des Klimawandels über die Gefahren für viele andere Küsten auf der Welt und die dort wohnenden Menschen im Klaren sein und entsprechend verantwortungsbewusst handeln. Nur dann können wir mit gutem Gewissen die Schönheit der Ostsee genießen.
Entwicklung der Ostsee
| Name | Leitfossil | Salzgehalt | Zeit |
| Baltischer Stausee | süß | 13.000-10.000 | |
| Yoldia Meer | Yoldia arctica | salzig | 10.000-9.500 |
| Ancylus See | Ancylus fluviatilis | süß | 9.500-8.000 |
| Litorina Meer | Littorina Littorea | salzig/brackig | 8.000-2.000 |
| Limnaea Meer | Limnea ovata | brackig | 2.000-500 |
| Mya Meer | Mya arenaria | brackig | 500-heute |

Wie entstehen Fossilien
Fotos und Text: Klaus Vöge

"Fossilien" ( latein. > fodere = graben ) war früher der Sammelbegriff für alle Dinge aus dem Erdreich, die ausgegraben wurden. Auch antike Reste. Der Begriff bezieht sich heute nur auf Versteinerungen von vorzeitlichen Tieren und Pflanzen. Das fossilführende Gestein (Sedimentgestein) ist ein Teil eines ehemaligen Meeres oder Festlands. In den Sedimentgesteinen ist meistens eine Vielzahl an unterschiedlichen Formen der Tier- und Pflanzengruppen enthalten. Dabei können einzelne Organismenreste in solchen Massen auftreten, dass sie gesteinsbildend sind.
Fossilbildung
Seit Jahrmillionen leben und sterben auf der Erdoberfläche eine Vielzahl von Pflanzen und Tieren, die ihre Spuren für uns hinterlassen haben und die wir zu deuten versuchen. Normalerweise werden die Lebewesen nach dem Tode in ihre ursprünglichen Bestandteile zurückverwandelt. Die Weichteile, die Knochen, die Panzer und die Schalen werden unter Mithilfe von Mikroorganismen chemisch zersetzt (verwesen) oder durch Wind und Wetter mechanisch zerrieben. Ein gefundenes Fossil ist immer ein Zufall der Natur. Das Lebewesen muss nach seinem Tode so bald wie möglich eingebettet werden (z.B. die Ameise im Bernstein, das Urpferd im Moor oder die Muschel im Schlamm) damit der Vorgang der Zersetzung nicht beginnen kann. Auf dem Festland bestehen die Voraussetzungen für den Versteinerungsprozess, bedingt durch die Witterung, nicht so häufig wie im Meer. Hier werden die Verwitterungsprodukte vom Land und die abgestorbenen Organismen des Meeres zusammengetragen und lagern sich als gesteinsbildendes Sediment ab. In diesen Sedimenten sind die Möglichkeiten der strukturerhaltenden Einbettung von toten Lebewesen erheblich höher. Deshalb werden maritime Fossilien häufiger gefunden als die von Landlebewesen. Trotzdem bleibt der Fossilbildung immer eine schöne Seltenheit.
Fossilerhaltung
Das Sediment (Schlamm, Sand, Kalkschlamm) wandelt sich im Laufe der Jahrmillionen in Gestein (Tone, Sandstein, Kalke) um. Die eingebetteten Organismen bzw. deren Hartteile nehmen an dieser Gesteinsbildung teil. Durch den entstehenden Druck werden die Lebewesen dabei jedoch zum Teil zerquetscht, flachgedrückt oder verformt. Es gibt viele Fossilien, an denen man die Kräfte erkennen kann, die auf sie eingewirkt haben. So sprechen wir von Ganzkörpererhaltung, wenn Schalenteil und Füllung erhalten ist. Eine Steinkernerhaltung liegt vor, wenn die Schale des Tieres weggelöst ist und nur noch die Füllung des Hohlraumes vorhanden ist. Sind sowohl die Schale und der Hohlkörper weggelöst, so nennt man die im Sediment vorhandene Negativform des Fossils eine Hohlformerhaltung. Weitere wichtige Faktoren bei der Fossilbildung sind chemische Umwandlungsprozesse in Verbindung mit Wasser. Nur ganz selten haben die Organismenreste in ihrer Ursprünglichkeit die Zeit überstanden. Das Wasser, das durch die Gesteinsporen dringt und Mineralstoffe mitführt, bewirkt, dass die eingeschlossenen Teile chemisch umgesetzt oder sogar chemisch völlig ausgetauscht werden. Die Flinte bzw. Feuersteine, die uns so häufig auffallen, sind ein Beispiel, wie ganze Sedimentschichten umkristallisiert werden können. Bei der Zersetzung von abgestorbenen Kieselalgen und Kieselschwämmen dringt die gelöste Säure (SiO2) in das Sediment und bildet hier die Feuersteinknollen. Durch diesen Prozess werden auch Fossilien im Feuerstein eingeschlossen oder selbst zu Flint umgewandelt. Derartige Vorgänge der Umkristallisation können sich im Laufe der Jahrmillionen mehrfach wiederholen.
Fossileinbettung 
Sie befasst sich mit der Beziehung eines Organismus und seiner Lage im umgebenden Sediment, wobei der Weg zwischen seinem Tod und der Lagerung nur schwer zu rekonstruieren ist. Im seltensten Fall entspricht der Todesort dem ehemaligen Lebensort.
Meist erfolgt die Einbettung jedoch nach einem längeren oder kürzeren Transportweg zu Lande oder im Wasser. Es ist verständlich, dass weite Transportwege die Fossilerhaltung nicht gerade begünstigen. Die Organismen werden z.T. in ihre Bestandteile zerlegt, auf dem Weg zerrieben oder anderweitig zerstört. So sind am Ort der endgültigen Einbettung nur noch Teile von den Skelettelementen vorhanden. Eine Rolle bei der Einbettung spielt auch die Wasserbewegung. So werden z.B. Muscheln fast immer mit der Wölbung nach oben eingebettet, da der Strömungswiderstand so am geringsten ist. Kegelförmige Organismen (Belemniten) werden gleichmäßig ausgerichtet.

Leitfossilien haben eine besondere Bedeutung innerhalb der Paläontologie. Diese Fossilien sind nur in einer geologisch kurzen Zeitspanne vertreten, jedoch über eine weite geographische Verbreitung, wodurch weit voneinander entfernte Gesteinsschichten einer Zeitspanne zugeordnet werden können. Diese grobe Zuordnung wurde durch genaue Beobachtung über die Jahrzehnte immer mehr verfeinert. Später wurde die Zeitdauer der einzelnen Perioden aufgrund des radioaktiven Zerfalls von Isotopen absolut festgelegt.
Faziesfossilien waren zu Lebzeiten an ein bestimmtes Milieu (Fazies) gebunden. Z.B. sind Korallen mit einem Riff verbunden und das Leben auf den Riffen ist miteinander vergleichbar.
Spurenfossilien sind Hinterlassenschaften ehemaliger Lebewesen wie Wohnbauten, Fressbauten, Weidespuren, Ruhe- und Kriechspuren. Sie können sowohl im Sediment selbst, wie auch in oder an Resten anderer Organismen auftreten. Die Verursacher der Spuren sind in den wenigsten Fällen bekannt.
Lebende Fossilien sind stammesgeschichtliche Lebewesen, die sich als einzelne Art über einen geologisch langen Zeitraum erhalten haben und heute noch leben.
Pseudofossilien sind Gebilde, die aussehen als seien sie echte Versteinerungen. Ihre Entstehung ist jedoch chemischer, physikalischer oder mechanischer Herkunft. Ein Beispiel sind die Dendriten (Eisen- und Manganoxyd) in den Solnhofener Platten, die an Pflanzen erinnern.
Müssen wir die rechtzeitige Einbettung eines Lebewesens schon als Sonderfall ansehen und berücksichtigen wir außerdem die Vielzahl der Zerstörungsmöglichkeiten, dann wird uns klar, dass nur häufig vorkommende Lebewesen als Fossil gefunden werden können und jeder Fossilienfund eine Seltenheit darstellt.
BELEMNITEN DER OBERKREIDE
von Carsten Rohde
Die Familie Belemnitellidae PAVLOW, 1914
|
Teil 3:
Artbegriff und Artenbestimmung |
Teil 3: Artbegriff und Artenbestimmung
Jede Artbestimmung ist zunächst eine Frage des subjektiven optischen Eindrucks. Sie ist nie wahr, sondern stets nur wahrscheinlich. Die verbale Beschreibung eines Rostrums ist durch die Wahl und die individuelle Interpretation der verwendeten, das Fossil beschreibenden Adjektive, wie klein/groß, kegel-/pfahl-/pflockförmig/zylindrisch oder etwa schlank/gedrungen, nicht widerspruchsfrei. Die Übergänge sind vielseitig und die Schattierungen zahlreich. So bezeichnet Jurij Alexander Jeletzky (1951, S.78f) die Rostrumform sowie die Ausprägung oder die Stärke der Rostrumskulptur der Belemnitellen und Belemnellen als zur Bestimmung vollkommen unzuverlässig. Deshalb ist bei den merkmalsarmen Resten der Belemnitelliden eine Hinwendung zur Inanspruchnahme der "inneren" Merkmale (Jugendrostren, Alveolentiefe, Apikallinie) sowie biometrischer Verhältniszahlen der gemessenen Merkmalswerte zu erkennen, die mit statistisch empirischen Methoden gewonnen, untersucht und, wie in nachstehender Tabelle 1 von Walter Kegel Christensen (2000, S. 60) dargestellt werden. Mittelwerte der gewählten Eigenschaften von betrachteten Populationen bleiben jedoch abstrakt. Nur das einzelne Individuum, das vermessene Rostrum ist real.
Tabelle 1 + Abbildung 1
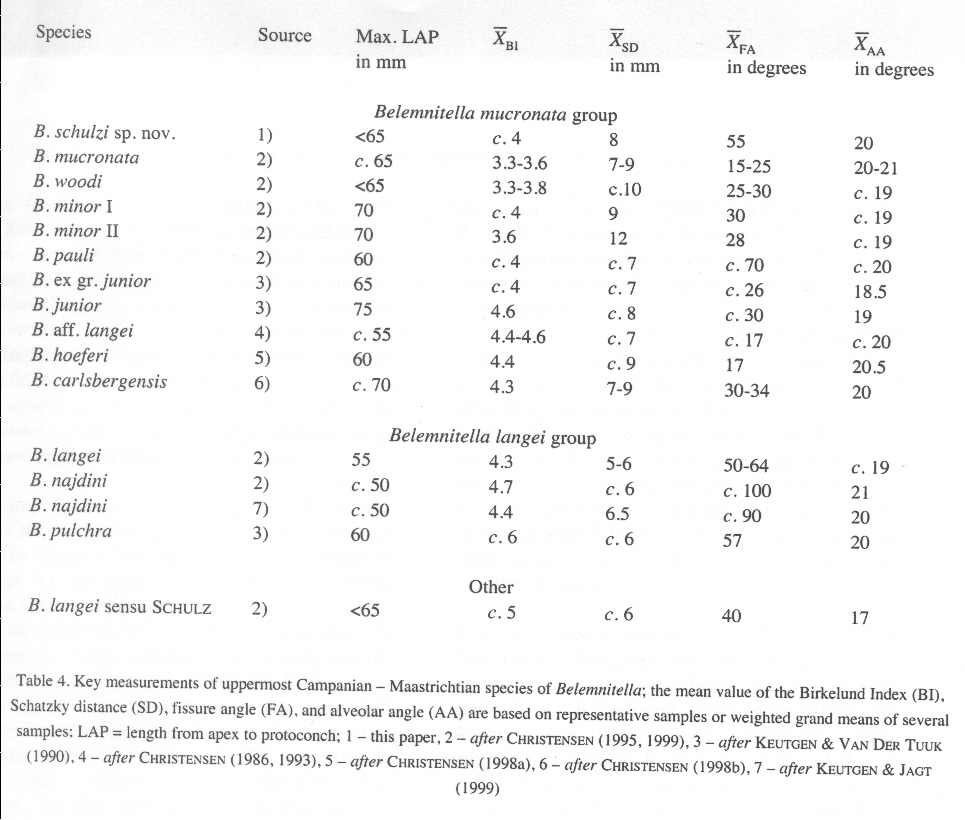
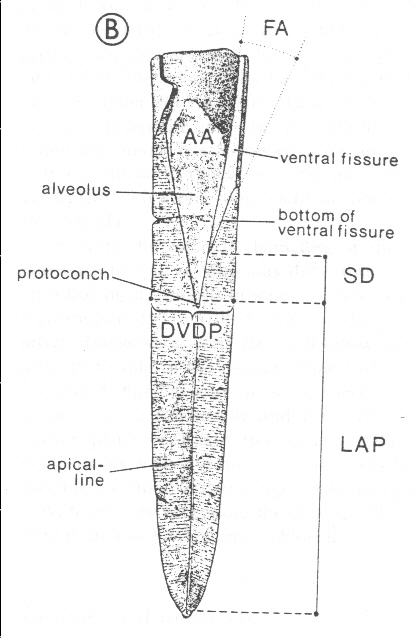 (LAP = Länge vom Apex bis zum Protoconch; BI = Birkelund-Index = LAP/Rostrumdicke in der Schlitzebene in Höhe des Protoconchs DVDP; SD = Schatzky-Wert = Entfernung vom Protoconch bis zur Einmündung des Ventralschlitzes in die Alveole; AA = Alveolarwinkel)
(LAP = Länge vom Apex bis zum Protoconch; BI = Birkelund-Index = LAP/Rostrumdicke in der Schlitzebene in Höhe des Protoconchs DVDP; SD = Schatzky-Wert = Entfernung vom Protoconch bis zur Einmündung des Ventralschlitzes in die Alveole; AA = Alveolarwinkel)
Die Wahl von Merkmalen führt immer zu typologischen Arten, sogenannten Morphospezies, die, im Gegensatz zu biologischen Arten, jederzeit aufgelöst und neu gegliedert werden können. Der paläontologische Artbegriff ist praktisch deshalb gleich dem morphologischen Artbegriff, da an Fossilien artkennzeichnende, biologische Merkmale zumeist nicht erhalten sind und die im Zeitquerschnitt geltende reproduktive Isolation nicht nachgewiesen werden kann (- wie kreuzt man zwei Rostren?). Ergebnis dieses Dilemmas: Jedwede vom jeweiligen Forscher als signifikant eingeschätzte Formabweichung erhält den Status eines artbestimmenden Merkmals; und im Zeitlängsschnitt werden Populationen aufeinander folgender geologischer Schichten solange als einer Art zugehörig betrachtet (einer sog. Chronospezies), wie ihre Merkmale nicht über einen statistisch als relevant betrachteten Bereich der empirisch erhobenen Merkmalswerte hinausreichen. Doch Veränderungen der Umwelt in der Zeit können das sichtbare Erscheinungsbild (den Phänotyp) folgender Generationen von Populationen beliebig stark wandeln, ohne zwangsläufig zur Umwandlung einer Art in eine andere neue Art (ein Wandel des Genotyps) zu führen, sondern lediglich zum Wechsel in den Merkmalsunterschieden zeitlich gestaffelter Populationen ein und derselben Spezies. Werden diese, durch die phänotypische Plastizität hervorgerufenen "Evolutions"stufen einer lückenlosen, sich nicht aufspaltenden Formenreihe als verschiedene Arten bezeichnet, so ist zunächst festzustellen, dass hierfür keine natürlichen Grenzen existieren. Diese Grenzen werden erst durch subjektive Gliederung der Formenreihe mittels empirischer Statistik geschaffen, sind damit im Sinne des biologischen Artkonzeptes willkürlich und definieren allenfalls mögliche, häufig auch sinnvolle Abgrenzungen auf dem Niveau von Unterarten.
Als eine gelungene zeitliche Aufspaltung einer phylogenetischen Entwicklungsreihe wird die Belemnitengattung Gonioteuthis BAYLE 1878 betrachtet. Ausgehend von Gonioteuthis praewestfalica ERNST & SCHULZ 1974 im späten Mittel-Coniac, über Gonioteuthis westfalica SCHLÜTER 1874 erfolgt eine lückenlose Entwicklungsreihe zu Gt. westfalicagranulata STOLLEY 1897 , Gt. granulata BLAINVILLE 1827, Gt. granulataquadrata STOLLEY 1897, Gt. quadrata quadrata BLAINVILLE 1827 bis zu Gt. quadrata gracilis STOLLEY 1892 im Oberen Unter-Campan. Die Abgrenzung der Gonioteuthisarten erfolgt anhand des Riedel-Quotienten, eines biometrischen Wertes, gebildet als Verhältnis von Rostrumlänge zu Alveolentiefe. Die Forschungen von Gundolf Ernst (1964, S. 169) zeigen Etappen der Stammesgeschichte der Gonioteuthiden mit allmählichen Übergängen und sich überschneidenden Abschnitten der Evolutionsprofile - Indizien für eine lückenlose, nicht verzweigte phylogenetische Entwicklung der Gattung Gonioteuthis. Mithin liegt der Verdacht nahe, dass es sich bei den Arten der Gattung Gonioteuthis um Chronospezies handelt. In Anwendung des biologischen Artbegriffs müssten daher alle Gonioteuthisarten als Unterarten eines polytypischen Stammes angesehen werden, der nach Blainville 1827 entweder mit Gonioteuthis granulata oder quadrata zu benennen wäre, zuzüglich eines differenzierenden Namens für die jeweilige Unterart.
Tabelle 2
Die Aufgabe empirischer Wissenschaft besteht darin, befriedigende Erklärungen zu finden, die nicht aus dem Augenblick heraus entstanden sind und überprüft werden können. Empirische Wissenschaft basiert dabei stets auf Stichproben mit endlicher Zahl von Individuen. Je größer die betrachtete Population, desto aussagefähiger die gewonnenen Ergebnisse. Stichproben mit nur wenigen Individuen können zu dubiosen Ergebnissen führen, zur Festlegung von Arten mit geringer Aussagefähigkeit und Akzeptanz.
Nach W. K. Christensen (2000,S.62) wurde die Belemnitellidenart Belemnitella posterior KONGIEL 1962 anhand von 7 nahezu vollständigen und 3 fragmentarischen Rostren im Übergang Obercampan/Untermaastricht begründet. Die mittleren statistischen Werte der von Christensen vermessenen Rostren liegen, wie nachstehende Tabelle zeigt, im selben Bereich wie die Werte der von Christensen neu etablierten Art Belemnitella schulzi CHRISTENSEN 2000 und sind der Art Belemnitella minor II CHRISTENSEN so ähnlich, dass er selbst "noted that it was uncertain if these were extreme variants of B. minor II or represented a separate species."
Da sich auch die stratigrafischen Reichweiten der vorgenannten Arten überlappen, erscheint eine Identität von posterior und schulzi nicht ausgeschlossen, wobei posterior der ältere Artname wäre. Und was hindert uns daran den Bogen größer zu schlagen und den Zweifeln Christensens folgend beide vorgenannten Arten als Varianten in Belemnitella minor II zu integrieren?
Doch auch bei hinreichend großer Zahl der untersuchten Rostren: Berechnungen anhand von Stichproben verschaffen keine Gewissheit, sondern nur Wahrscheinlichkeit. So sind stark vom Mittelwert abweichende Ergebnisse in der Paläontologie nicht zwingend Ausdruck von Pathologie und damit zu eliminieren; Abweichler können auch Ausdruck einer hohen Variabilität der Art sein, d.h. auf ein großes Potenzial an Variantenbildung hinweisen. Sie können aber auch schlicht eine neue Art angeben. Doch wo hört die Variabilität einer Art, ihre phänotypische Plastizität auf, wo wird die Normalität der einen Art zu der Normalität ihrer Schwesterart? Es gibt innerhalb der Belemnitengattungen durchaus Arten hoher stratigraphischer Reichweite und grosser Variabilität , wie Hibolites jaculoides SWINNERTON 1936 (Ober-Valangin Oberstes Barrême), neben solchen mit kurzer Lebensdauer und unveränderlichen Merkmalsausprägungen (Belemnellocamax boweri CRICK 1910, frühes Mittel-Cenoman).
Weitere Schwierigkeiten bei der Artabgrenzung ergeben sich aus den Problemen der Formähnlichkeit (Homöomorphie, so tauchen in der Entwicklungsgeschichte der Belemniten zu verschiedenen Zeiten kleine, keulenförmige Rostren auf, die kaum zu unterscheiden sind), der Heterogenität zwischen Individuen unterschiedlichen Alters, durch ausgeprägten Geschlechtsdimorphismus (von Schulz bei den Belemnellen angenommen, von Peter Doyle bei Youngibelus aus dem Toarc von Yorkshire beschrieben) und pathologische Veränderungen an Rostren gleicher Artzugehörigkeit. Zudem sind die Individuen einer Art in ihren Merkmalsausprägungen nie deckungsgleich; wie auch Populationen gleicher Art an verschiedenen Standorten, hervorgerufen durch die abweichenden Milieubedingungen, möglicherweise erhebliche Merkmalsunterschiede aufweisen werden.
Bei der Bestimmung von Arten der Belemnitenfamilie Belemnitellidae werden verschiedene biometrische Verhältniszahlen (Riedel-Quotient, Birkelund-Index) oder gar von komplexen Merkmalskombinationen (bei der Abgrenzung der Belemnellen) gebildet. Derartige Verhältniszahlen sind aber nur dann geeignet Arten zu differenzieren, wenn ihre Schwankungen nur Ausdruck der Normabweichung sind und nicht durch Messfehler oder unterschiedliches Wachstumsverhalten im Alterungsprozess beeinflusst wird.
Der gezielte Einsatz der Variationsstatistik und biometrischer Messmethoden bei der Bestimmung von Belemnitenrostren kann als Mittel dienen, eine Auflösung des Artbegriffs durch unterschiedliche Interpretation beschreibender, qualitativer Merkmale zu verhindern. Sie kann aber auch das Gegenteil bewirken, da sie nicht wie die altmodische subjektive Deutung von der Gesamtheit der beobachteten Merkmale ausgeht, sondern nur von einigen biometrisch erfassbaren. Auch sollte die Variabilität der Arten nicht zu gering geachtet werden, wie viele Beispiele aus der Neontologie zeigen.
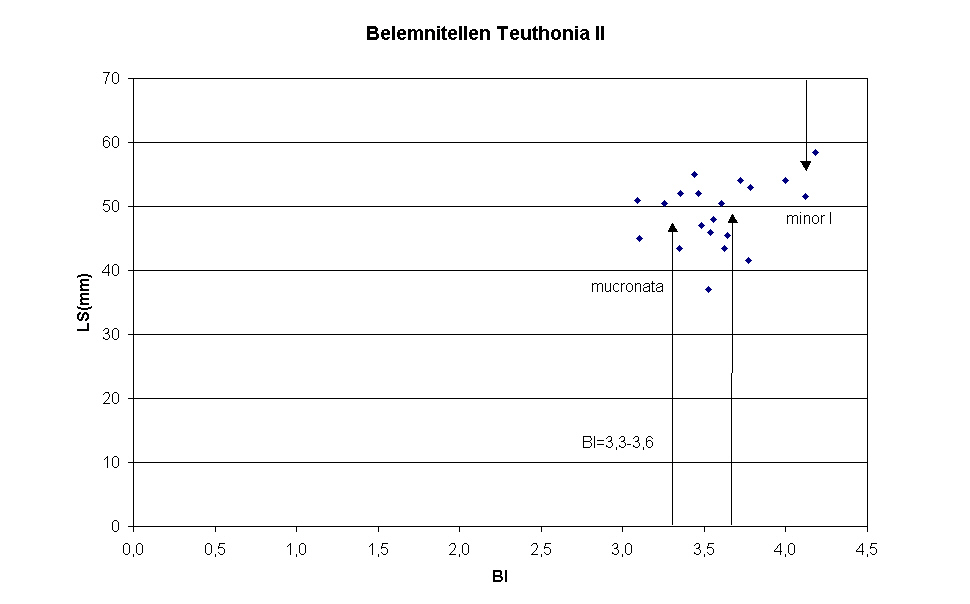
Im Jahre 2005 wurden in der Grube Teutonia Nord der Teutonia AG in Misburg (bei Hannover), die stratigraphisch von der pilula/senonensis-Zone des Unter-Campan bis zur minor/polyplocum-Zone des Unteren Ober-Campan reicht, 20 Belemnitenrostren nicht horizontiert gesammelt. Nach Reinigung der Rostren mit Hilfe des Tensids Rewoquat®, wurden anhand der Alveole geprüft, ob die Rostren zu der Gattungen Gonioteuthis BAYLE 1879 (kurze Pseudoalveole mit wenige mm langem Alveolarschlitz und wulstigem Alveolarrand) oder Belemnitella dORBIGNY 1840 (ausgeprägte Alveole und langer Alveolarschlitz) gehören. Alle Rostren konnten der Gattung Belemnitella zugeordnet werden. Zur Bestimmung der Lage des Protoconch wurde die Alveole, soweit möglich, aufgebohrt; bei den meisten Rostren war indes das eingeschlossene Sediment zu fest, um es zu entfernen. Diese Exemplare wurden in Höhe der vermuteten Lage der Embryonalkammer quergeteilt. Aufgrund der nunmehr geringeren Alveolentiefe konnte die Position des Protoconchs durch Entfernen des geringen verbliebenen Sediments leicht bestimmt werden. Lag der Bruch Apex-seitig unterhalb der Embryonalkammer, wurde das alveolenseitige Rostrumfragment in der Alveolarschlitzebene gespalten. Die Werte für die Rostrumlänge bis zu Protoconch LAP sowie der Birkelund-Index BI wurden in Diagramm 1 abgetragen. Das Schaubild zeigt, dass der Birkelund-Index der meisten Rostren innerhalb des von Christensen in Tabelle 1 angegebenen Bereichs von 3,3 bis 3,6 liegt, der Belemnitella mucronata LINK 1807 repräsentiert. Die BI- Werte der Rostren 1, 2 und 12 entsprechen dem Mittelwert des BI-Index für Belemnitella minor JELETZKY 1951 in der von Christensen eingeführten Variante minor I.
|
Analyse III:
Belemnitella Arten aus der Grundgesamtheit von 20 nicht Horizont orientiert gesammelten Rostren in Teutonia Nord/Misburg
|
|||
|
Rostrum
|
LS(LAP)
|
DP
|
BI
|
| 1 | 54 | 13,5 | 4,0 |
| 2 | 58,5 | 14 | 4,2 |
| 3 | 53 | 14 | 3,8 |
| 4 | 46 | 13 | 3,5 |
| 5 | 45 | 14,5 | 3,1 |
| 6 | 51 | 16,5 | 3,1 |
| 7 | 45,5 | 12,5 | 3,6 |
| 8 | 41,5 | 11 | 3,8 |
| 9 | 43,5 | 12 | 3,6 |
| 10 | 47 | 13,5 | 3,5 |
| 11 | 52 | 15 | 3,5 |
| 12 | 51,5 | 12,5 | 4,1 |
| 13 | 54 | 14,5 | 3,7 |
| 14 | 50,5 | 14 | 3,6 |
| 15 | 48 | 13,5 | 3,6 |
| 16 | 50,5 | 15,5 | 3,3 |
| 17 | 55 | 16 | 3,4 |
| 18 | 52 | 15,5 | 3,4 |
| 19 | 37 | 10,5 | 3,5 |
| 20 | 43,5 | 13 | 3,3 |
| 21 | 51 | 15,5 | 3,3 |
| 22 | 53 | 14 | 3,8 |
|
STABW
|
0,3
|
||
|
Mittelwert
|
3,6
|
||
Literaturhinweise:
Christensen, W.K. (1997): The Late Cretaceous belemnite family Belemnitellidae: Taxonomy and evolutionary history. Bulletin of the Geol. Soc. Denmark, Vol. 44, S. 59-88, Kopenhagen.
Christensen, W.K. (2000): Belemnitella schulzi sp.nov. from the uppermost Campanian and lowest Maastrichtian chalks of northwest Germany and Denmark. Acta Geologica Polonica, Vol. 50, S. 55 66, Warschau.
Ernst, G. (1964): Ontogenie, Phylogenie und Stratigraphie der Belemnitengattung Gonioteuthis BAYLE aus dem nordwestdeutschen Santon/Campan. Fortschr. Geol. Rheinld. u. Westf. 7, Krefeld.
Jeletzky, J.A. (1951): Die Stratigraphie und Belemnitenfauna des Obercampan und Maastricht Westfalens, Nordwestdeutschlands und Dänemarks sowie einige allgemeine Gliederungs-Probleme der jüngeren borealen Oberkreide Eurasiens. Beih.Geol.Jb. 1, Hannover.
Mayr. E. (1975): Grundlagen der zoologischen Systematik, Hamburg/Berlin.
Schulz, M.-G. (1979). Morphometrisch-variationsstatistische Untersuchungen zur Phylogenie der Belemnitengattung Belemnella im Untermaastricht NW-Europas, Geol.Jb. A47, Hannover.
|
Teil 3:
Artbegriff und Artenbestimmung |
Wie der „Alte Schwede“ ein Hamburger wurde
von Klaus Vöge
 Seit 15 Jahren ist Hamburg um eine schwergewichtige Sehenswürdigkeit reicher. Bei Baggerarbeiten zur Elbvertiefung wurde ein gewaltiger Findling in der Elbe entdeckt. In der Elbe konnte er nicht bleiben. Also was sollte mit diesem Brocken passieren? Sprengen oder Bergen. Die Entscheidung fiel zu Gunsten des Findlings aus und er sollte geborgen werden. Aber ganz so einfach machte es der Stein dem Schwimmkran dann doch nicht. Beim ersten Versuch zeigte sich der Findling ca. 1 ½ Meter über der Wasseroberfläche, rutschte aus der Halterung und versank wieder in der Elbe. Bei diesem Versuch zeigte sich aber auch, dass der Stein um etliches schwerer war als er ursprünglich geschätzt wurde. Der zweite Bergungsversuch verlief dann erfolgreich. Der Stein wurde auf eine Schute verladen und an den Strand von Övelgönne transportiert, wo er an einem vorbereiteten Platz aufgestellt wurde.
Seit 15 Jahren ist Hamburg um eine schwergewichtige Sehenswürdigkeit reicher. Bei Baggerarbeiten zur Elbvertiefung wurde ein gewaltiger Findling in der Elbe entdeckt. In der Elbe konnte er nicht bleiben. Also was sollte mit diesem Brocken passieren? Sprengen oder Bergen. Die Entscheidung fiel zu Gunsten des Findlings aus und er sollte geborgen werden. Aber ganz so einfach machte es der Stein dem Schwimmkran dann doch nicht. Beim ersten Versuch zeigte sich der Findling ca. 1 ½ Meter über der Wasseroberfläche, rutschte aus der Halterung und versank wieder in der Elbe. Bei diesem Versuch zeigte sich aber auch, dass der Stein um etliches schwerer war als er ursprünglich geschätzt wurde. Der zweite Bergungsversuch verlief dann erfolgreich. Der Stein wurde auf eine Schute verladen und an den Strand von Övelgönne transportiert, wo er an einem vorbereiteten Platz aufgestellt wurde.
Hier ist er zu einem richtigen Fotomodell geworden. Leider wird er auch regelmäßig von Sprayern aufgesucht. Nachdem er seinen festen Platz erhalten hatte, wurde er wissenschaftlich untersucht, um die anfallenden Fragen zu beantworten. Als erstes konnte das Gewicht ermittelt werden; das war schnell möglich, denn der Schwimmkran hat eine eingebaute Waage. Es sind stolze 217 Tonnen Gestein. Sein Umfang beträgt fast 20 Meter, seine Höhe 4,5 Meter und die Breite zwischen 8,0 und 5,2 Meter. Damit ist dieser Fund aus der Elbe einer der größten Findlinge im norddeutschen Raum. Die Presse machte ihn zum größten Findling Europas, aber da sind noch einige Findlinge gewaltiger (z.B. der Große Stein von Altentreptow mit 360 Tonnen und der Findling „Buskam“ vor Rügen mit einem geschätzten unglaublichen Gewicht von 1.600 Tonnen). Dafür hat der Hamburger „Zugereiste“ den schönsten Ausblick auf die vorbeifahrenden Schiffe auf der Elbe.
Apropos „Zugereister“: um welches Material handelt es sich, wie alt ist unser Stein, wann ist er zu uns eingereist und wie ist er zu uns gekommen? Diese Fragen haben uns Herr Prof. R. Vinx und Herr Dr. J. Ehlers beantwortet. Sie befassen sich mit diesen Themen wissenschaftlich. Der Granit kann im engsten Sinne als grauer Ostsmålandgranit aus der Region von Växjö (oder Växjögranit) angesehen werden und ist rund 1,8 Milliarden Jahren alt. Für den Transport zu uns sind die Gletscher der letzten drei Kaltzeiten verantwortlich. Die Gestaltung der Landschaft von Schleswig-Holstein ist das Werk von Kaltzeiten und den jeweils folgenden Warmzeiten. Die Gletscher haben in den Jahrtausenden eine riesige Menge von Sand und Steinen (Geschiebe) aus dem skandinavischen Raum zu uns transportiert. Hier die letzten drei nördlichen Kaltzeiten:
 Weichselkaltzeit vor 80.000 – 15.000 Jahren
Weichselkaltzeit vor 80.000 – 15.000 Jahren
Saalekaltzeit vor 200.000 – 125.000 Jahren
Elsterkaltzeit vor 300.000 – 250.000 Jahren
Ältere Kaltzeiten sind bis heute in Norddeutschland nicht nachgewiesen.
Da die Gletscher der Weichselkaltzeit die Elbe nicht überschritten haben, kommen nur die beiden älteren Gletschervorstöße für den Transport infrage, die sich weiter in Richtung Westen ausgebreitet haben. Bei der Zeitbestimmung des Transports, kommt ein weiterer Findling (der Stein von Othmarschen) ins Spiel. Dieser 60 Tonnen schwere Findling ist bei den Arbeiten zur vierten Elbtunnelröhre gefunden worden. Anhand des ihn umgebenden Geschiebematerials konnte nachgewiesen werden, dass dieser Stein von den Gletschern der Saalekaltzeit zu uns transportiert wurde. Bei unserem Findling von Övelgönne wurde anhand des umgebenden Materials festgestellt, dass der riesige Brocken mit der älteren Elsterkaltzeit vor 300.000 -250.000 Jahren zu uns gekommen ist. Nach dieser gewaltigen Zeitspanne wurde es auch Zeit, ihm endlich einen Namen zu geben. Der große Stein wurde am 6. Juni 2000 feierlich auf den Namen „Alter Schwede“ getauft und damit wurde er offiziell eingebürgert.
Abschließend möchte ich noch kurz beschreiben wie die Landschaft Schleswig-Holsteins durch die Eiszeiten der letzten 300.000 Jahre geprägt wurden. Es ist kaum vorstellbar, aber es gab in der jüngeren Erdgeschichte Zeiten in denen große Teile Nordeuropas über mehrere tausend Jahre unter einer mächtigen Eisdecke begraben waren. Infolge der Abkühlung kam es in Nordeuropa verstärkt zu Schneefällen. Es gab viel mehr Schnee als in den Sommermonaten abschmelzen konnte. Dadurch häufte sich der Schnee gewaltig an und wurde durch das Eigengewicht zu Eis zusammengepresst. In den Nährgebieten wuchs das Eis bis zu einer Höhe von 3.000 bis 5.000 Meter. Wie viel Schnee muss in den Bergen Skandinaviens gefallen sein, wenn für 1 Meter Eis 80 Meter Schnee fallen muss. Da Eis plastisch ist, begannen die gewaltigen Eismassen vorzurücken. Die Gletscher flossen über hunderte Kilometer von der norwegischen Hochplateaufläche in südliche und südwestliche Richtung. Das Eis übte eine ungeheure Kraft auf den Untergrund und die Flanken aus. Gestein in unterschiedlicher Größe wurde losgerissen, Felsen zermahlen und mitgeführt. Das sogenannte „Geschiebe“ luden die Gletscher nach einer langen Reise bei uns im Norddeutschen Raum ab.
Welche Mengen da bewegt wurden, erahnt man, wenn man weiß, dass wir in Schleswig-Holstein auf einer Geschiebeschicht von 200 bis 400 Metern leben. Charakteristische Hinterlassenschaften der Gletscher sind neben Moränen und Sanderflächen auch die Gesteinsstrände der Ostsee mit ihren Geschiebemergelkliffs. Kennzeichnend ist die Durchmischung aller Korngrößen vom feinsten Partikel bis zum Findling. Ein weiteres Charakteristikum sind die Gletscherschrammen oder sogenanntes Gekritztes, das durch gegenseitiges Reiben der Gesteine während des Transports entstanden ist. Wenn wir bedenken, dass das Gletschereis in Schleswig-Holstein noch eine Stärke von 300 bis 500 Meter hatte, ist verständlich, dass der „Alte Schwede“ kein Transportproblem darstellte. Da die Gletscher in Skandinavien auch die Sedimentgesteine abgebaut und zu uns transportiert haben, können wir in den Kiesgruben und an der Küste die von uns gesuchten Fossilien finden, zwar nicht sortiert, dafür aus allen Erdzeitaltern.
(Bilder anklicken, um die Galerie zu öffnen.)





")



Belemniten der Oberkreide
von Carsten Rohde
Die Familie Belemnitellidae PAVLOW, 1914
|
Teil 2:
Entwicklungsgeschichtliche Zusammenhänge |
Teil 2: Entwicklungsgeschichtliche Zusammenhänge
Wichtige frühe Forschungsarbeiten zur Entwicklungsgeschichte der Familie Belemnitellidae stammen von dem russischen Forscher Dmitrij Pavlovic Najdin und dem viele Jahre in Kanada lebenden russischen Wissenschaftler Jurij Alexander Jeletzky (1915, 1988). In jüngerer Zeit wurden Gattungszusammenhänge und Ursprung von Walter Kegel Christensen (1942, 2002) in vielen Artikeln beschrieben und teilweise neu geordnet.
Die Zusammenfassung verschiedener Belemnitengattungen der Oberkreide zu der Familie der Belemnitellidae führte Pavlow 1914 ein. Einige dieser Gattungen wurden dagegen schon erheblich früher bestimmt: Actinocamax durch MILLER 1823 und Belemnitella durch d`ORBIGNY 1840. Die moderne Belemnitenforschung hat insbesondere seit den 60er Jahren mit statistisch-mathematischen Analyseverfahren, den sogenannten biometrischen Methoden, neue Zuordnungen definiert und neue Gattungen kreiert: So etwa Belemnella (Pachybelemnella) durch SCHULZ 1979 und Praeactinocamax durch Košták 2004.
Bis zum Jahre 1964 wurde die Familie der Belemnitelliden in fünf Gattungen gegliedert: Actinocamax, Belemnella, Belemnitella, Belemnocamax und Gonioteuthis. Najdin (1964) erweiterte diese Systematik durch eine neue Klassifikation der Rostren mit flachem, konvexem alveolarseitigem Rostrumstumpf oder mit einer Pseudoalveole; letztere entsteht in der Regel durch Verwitterung der aragonitischen Teile des Alveolenrandes im Sediment und ist, wie die Fotos im Anhang zeigen, bei den Hibolithen schon seit dem Oberen Malm bekannt und setzt sich bei ihren Nachfahren, der Familie Mesohibolitidae NERODENKO 1983, fort. Erst bei den späten Belemnitelliden des Obercampan und Maastricht wird der Alveolenrand calcitisch und damit erhaltungsfähig.
Najdin fügte den bekannten Belemnitelliden-Gattungen die Gattung Belemnellocamax hinzu, erweiterte Actinocamax um zwei Untergattungen: Praeactinocamax, Paractinocamax und ergänzte Gonioteuthis um die Untergattung Goniocamax. Christensen (1997) erhob die vorgenannten Untergattungen in den Rang von Gattungen und vereinte Paractinocamax mit Belemnellocamax.
Viele der neuen Gattungen waren in der Zentral Europäischen Subprovinz der Oberkreide unbekannt. Sie wurden erstmals in der Zentral Russischen Subprovinz erfasst oder in der Baltisch-Skandinavischen Region, in der sich beide Subprovinzen überlappen (siehe hierzu Teil 1). Nunmehr umfasst die Familie Belemnitellidae neun Gattungen und zwei Untergattungen und stellt sich, gegliedert nach der erstmaligen zeitlichen Bestimmung der Gattungen, wie folgt dar:
|
Gattungen
|
Anzahl der Arten
|
|
Actinocamax MILLER, 1823
|
4 |
|
Belemnitella dORBIGNY, 1840
|
17 |
|
Gonioteuthis BAYLE, 1878
|
9 |
|
Belemnocamax CRICK, 1910
|
1 |
|
Belemnella NOWAK, 1913
|
12 |
|
Fusiteuthis KONGIEL, 1962
|
1 |
|
Praeactinocamax NAJDIN, 1964
|
10 |
|
Belemnellocamax NAJDIN, 1964
|
3 |
|
Goniocamax NAJDIN, 1964
|
4 |
Über den Ursprung der Belemnitelliden gibt es noch keine Klarheit. Jeletzky (1946) nahm an, dass sie von den Neohiboliten des Apt (N. ewaldi STROMBECK 1861 und N. clava STOLLEY 1911) abstammen, da die äußere Form des Rostrums sowie die Form der Pseudoalveole sehr ähnlich sind. Dagegen vermutete Doyle (1992) einen ursprünglichen Zusammenhang mit den Hibolithen des Hauterive bis Apt (z.B. H. jaculoides SWINNERTON 1952). Beide Ansätze verkennen, dass zwischen dem Verschwinden der Unterkreide Hibolithen bzw. Neohiboliten und dem erstmaligen Auftreten von Praeactinocamax primus im Unteren Cenoman ein zeitliches „Loch“ von rund 20 Mio. Jahren klafft. Daher ist im Prinzip Najdin & Alekseev (1977) zu folgen, die eine zeitnahe Abstammung der Belemnitelliden von Unter-Cenoman Neohiboliten vorschlagen. Ihr „Übergangskandidat“, Neohibolites repentinus NAJDIN & ALEKSEEV 1975 kommt als Ahn jedoch nicht in Frage, da er nach Christensen (1997) später erschien, als sein vermeintlicher Nachfolger Praeactinocamax primus.
Steht der Ursprungsbeweis mithin noch aus, so ist auch die Abstammungslinie der Belemnitelliden keineswegs stimmig oder gar durchgehend, sondern vielmehr in weiten Bereichen noch ungeklärt.
Najdin & Kopaevich (1977: The zonation of the Upper Cretaceous of the European palaeobiogeographical province ( in russisch) zeigen stammesgeschichtliche Zusammenhänge zwischen den Gattungen der Belemnitelliden auf, ausgehend von der Gattung Praeactinocamax:
|
1. Praeactinocamax
|
führt zu Actinocamax
|
||
|
2. Praeactinocamax
|
über Belemnellocamax
|
führt zu Belemnella
|
|
|
3. Praeactinocamax
|
über Goniocamax
|
führt zu Gonioteuthis
|
führt zu Belemnitella
|
Die Annahme der Entwicklung der Belemnitelliden aus einer Ursprungsgattung ist umstritten. So sind sicher Praeactinocamax und Belemnocamax aufgrund ihres parallelen Auftretens und ihrer abweichenden morphologischen Merkmale nicht zeitlich unmittelbar ursprungsgleich. Zweifelhaft ist auch die Ableitung von Actinocamax aus Praeactinocamax, da beide nicht nur im Wachstumsverhalten (Actinocamax zeigt isometrisches, Praeactinocamax dagegen allometrisches Wachstum des Rostrums) von einander abweichen. Košták (2004) vermutet einen gemeinsamen Vorfahren beider Gattungen bei einem noch unbekannten Neohiboliten des Unter-Cenoman.
Die Familie Belemnitellidae ist allem Anschein nach ein polyphyletisches Taxon mit verschiedenen Ahnen: Denn obwohl die Wurzeln von Actinocamax und Praeactinocamax wohl in beiden Fällen bei späten Neohiboliten zu suchen sind, passt doch die kurze gedrungene Gestalt von Belemnocamax boweri CRICK 1910 mit einer tiefen, breiten Ventralfurche so gar nicht in das Bild der Belemnitelliden; wäre da nicht als Gemeinsamkeit die Pseudoalveole des Rostrums. Belemnocamax ausklammernd könnte man mit Košták durchaus korrekt von einer monophyletischen Entwicklung der Belemnitellidae sprechen.
Auch der häufig vermuteten Abstammung der Belemnellen von Belemnellocamax steht eine erhebliche stratigraphische Lücke entgegen, die sich zwischen dem Aussterben der einen Gattung im Unteren Obercampan und dem Erscheinen der Belemnellen im Unter-Maastricht auftut (ca. 9 Mio. Jahre).
Über die Zuordnung der wie Belemnocamax sehr seltenen Gattung Fusiteuthis herrscht Unklarheit.
Christensen (1997; Grafik 1) definiert die Hauptstammlinien der Familie Belemnitellidae daher wie folgt neu:
Grafik Christensen:

Grafik 1: Christensen, The Late Cretaceous belemnite family Belemnitellidae...
Bulletin of the Geol. Soc. of Denmark, Vol. 44, S.61
Wohl definiert (Grafik 2) und fossil belegt ist zur Zeit nur die Entwicklungslinie von Praeactinocamax plenus ausgehend über Goniocamax lundgreni zu den Gonioteuthiden und Belemnitellen des Santon.

Grafik 2: Christensen/Schulz, Coniacian and Santonian belemnite faunas... . Fossils and Strata, Nr. 44, S.20
|
Belemnocamax?
|
führt zu Actinocamax
|
||
|
Praeactinocamax
|
führt zu Belemnellocamax
|
||
|
Praeactinocamax
|
über ?Belemnellocamax
|
führt zu Belemnella
|
|
|
Praeactinocamax
|
über Goniocamax
|
führt zu Gonioteuthis
|
führt zu Belemnitella
|
|
?Praeactinocamax
|
über ?Belemnellocamax/?Belemnitella
|
führt zu Fusiteuthis
|
 Neohibolites ewaldi, Unter-Apt, Helgoland Neohibolites ewaldi, Unter-Apt, Helgoland |
 Neohibolites jaculoides, Hauterive; Moorberg; gr. Rostrum Neohibolites jaculoides, Hauterive; Moorberg; gr. Rostrum |
 Hibolites Minutus, mittleres, Barreme, Hoheneggelsen Hibolites Minutus, mittleres, Barreme, Hoheneggelsen |
 Neohibolites minimus, Apt, Soleilhas Castillon, F Neohibolites minimus, Apt, Soleilhas Castillon, F |
 Hibololites calloveyensis, Mittl. Callov, Nikitino, Rjazan Hibololites calloveyensis, Mittl. Callov, Nikitino, Rjazan |
 Praeactinocamax plenus, oberes Cenoman, Mühlheim-Broice Praeactinocamax plenus, oberes Cenoman, Mühlheim-Broice |
 Praeactinocamax plenus, Pseudoalveole Praeactinocamax plenus, Pseudoalveole |
 Goniocamax lundgreni + Gx striatus, Bavnodde, Grünsand Goniocamax lundgreni + Gx striatus, Bavnodde, Grünsand |
 Goniocamax lundgreni, Pseudoalveole |
 Actinocamax verus, Bavnodde, Grünsand, Coniac-Santon Bornh. |
Literaturhinweise:
Birkelund, T. (1957): Upper Cretaceous belemnites of Denmark. Det Kong. Danske Vidensk. Selskab 9, S. 1 69.
Christensen,W.K. (2002): Fusiteuthis polonica, a rare and unusual belemnite from the Maastricht. Acta Palaeontologica Polonica 47(4), S. 679 - 683.
Christensen, W.K., Schulz, M.-G. (1997): Coniacian and Santonian belemnite faunas from Bornholm, Denmark. Fossils and Strata, Nr. 44, S. 1 - 73, Oslo.
Doyle, P. (1992): A review of the Biogeography of Cretaceous belemnites. Palaeogeography, Palaeoclimatology, Palaeoecology 92, S. 207 - 216.
Košták, M. (2004): Cenomanian through the Lowermost Coniacian Belemnitellidae Pavlow (Belemnitida, Coleoidea) of the East European Province. GEOLINES 18, Prag, S. 59ff.
Košták, M. (2005): Phylogenie of Praeactinocamax NAIDIN (Belemnitellidae, Upper Cretaceous). 2nd Int. Symp.-Coleoid Cephalopods Through Time, Prag, S.80 - 84.
Najdin, D.P., Alekseev, A.S. (1975): New Neohibolites from Cenomanian of the Crimea. Übersetzung in: Palaeont. Journal 3, S. 311 - 315.
|
Teil 2:
Entwicklungsgeschichtliche Zusammenhänge |